私たちはナルディライジンという分子の研究を通して、様々な生命現象や疾患メカニズムの解明に取り組んでいます。ナルディライジンは、もともとグループリーダー(西 英一郎)が増殖因子の受容体として同定したメタロプロテアーゼの1種でしたが、その後の研究から、細胞外ドメインシェディングを増強する機能、核内での転写制御機能など、局在に応じた多機能性を有するユニークなタンパク質であることがわかってきました。ここでは、我々の研究の歴史と進行中のプロジェクトについて概説します。
目次
1)ナルディライジンの発見
2)ナルディライジンの細胞外ドメインシェディング活性化機能
3)ナルディライジン(NRDC)の生物学的機能と病態生理学的意義
3)-1:神経系発達におけるNRDCの役割
3)-2:アルツハイマー病におけるNRDCの役割
3)-3:体温恒常性維持機構におけるNRDCの役割
3)-4:糖尿病、メタボリック症候群におけるNRDCの役割
3)-5:循環動態制御におけるNRDCの役割
3)-6:心疾患におけるNRDCの役割
3)-7:がんにおけるNRDCの役割
3)-8:炎症性疾患におけるNRDCの役割
4)ナルディライジンの多機能性
5)おわりに
1) ナルディライジンの発見
グループリーダーは、京都大学老年科大学院(注1)で、動脈硬化におけるTリンパ球の役割に関する研究を行い、その過程でヘパリン結合性EGF様増殖因子(HB-EGF)という増殖因子の仕事をしました。その縁で、HB-EGFを精製同定した研究室に留学(注2)することになり、HB-EGFの新規受容体同定を目標に実験を開始しました。HB-EGFの既知のシグナリング受容体はEGF受容体(EGFR/erbB1)でしたが、EGFRには少なくとも7種類以上のEGFファミリー増殖因子が結合することがわかっていたため、HB-EGFの特異性がEGFR以外の共役受容体的分子で説明できる可能性があるのではないかと考えたからです。
放射性ヨード標識したHB-EGFを、様々な細胞表面にクロスリンクさせてみたところ、神経細胞や乳がん細胞表面で、EGFRとの複合体とは明らかに分子量の異なる複合体が検出されました。この複合体はEGFRやそのファミリー分子に対する抗体では免疫沈降できなかったことから、新規受容体との複合体である可能性があると考え、チューブ内クロスリンクによる複合体形成を指標として、タンパク質生化学的に精製を試みました。その結果、ナルディライジンの同定に至ったのですが、細胞表面上でHB-EGFと特異的に結合する分子として同定したにもかかわらず、ナルディライジンは膜貫通ドメインのない可溶型のメタロエンドペプチダーゼ(図1)でした。ナルディライジンに明らかなシグナル配列はなく、主に細胞質に存在しますが、非古典的分泌経路を通って細胞外にも分泌され、一部は細胞表面に留まります。この細胞表面のナルディライジンがHB-EGFと強固に結合していたのです
(Nishi他 EMBO.J, 2001)。
注1:北徹先生(現神戸医療センター中央市民病院院長)、久米典昭先生(現神戸学院大学薬学部教授)に師事。
注2:ハーバード大学小児病院 Michael Klagsbrun研究室:当研究室留学中にHB-EGFを同定された東山繁樹先生(現愛媛大学医学部教授)にご紹介頂きました。
(図1)

2) ナルディライジン(nardilysin, N-arginine dibasic convertase, gene nameおよび略称:NRDC)の細胞外ドメインシェディング活性化機能
膜近傍部でのタンパク分解により、膜タンパク質の細胞外ドメインが不可逆的に切断される現象を、細胞外ドメインシェディング(以下シェディングと略します)といいます。増殖因子やサイトカインの前駆体、様々な受容体や接着分子、さらにNotchやアミロイド前駆体蛋白質(APP)など、多岐にわたる膜タンパク質がシェディングによる制御を受けており、ADAM
(a disintegrin and metalloprotease) プロテアーゼをはじめとして、MMPファミリー分子やBACE1など様々な酵素群が“シェデース“(切断酵素)として報告されています。
HB-EGFを含むEGFファミリー増殖因子は全て、膜タンパク質前駆体として産生され、シェディングを受けて分泌型(活性型)になります。もともとNRDCは分泌型HB-EGFの受容体として同定したのですが、その後の研究から、HB-EGFの膜タンパク質前駆体にも結合し、さらにそのシェディングを増強することが明らかになりました(Nishi他 JBC,
2006)。NRDC自身もプロテアーゼ活性を有しますが、シェディング増強効果は同プロテアーゼ活性に依存せず、ADAMプロテアーゼの活性化を介して発揮されることがわかりました。さらに、NRDCのシェディング増強効果はHB-EGFに限定されず、アミロイド前前駆体蛋白質、TNF-αなど広範な膜タンパク質に及ぶことがわかりました(Nishi他JBC,2006、Hiraoka他J.Neurochem,2007、Hiraoka,
Yoshida他BBRC,2008)。
私たちの実験結果をもとに、シェディングの活性化における『NRDC-ADAMsシステム仮説』、すなわち、シェディング誘導因子の刺激により①『NRDCの細胞膜上への移動』が起こり,②『ADAMsと結合』することで、③『ADAMsのコンフォメーション変化による酵素活性の増強』を起こし,その結果④『基質膜蛋白質の細胞外ドメインシェディングの増強』を誘導するというシステム仮説を提唱しています(図2)。
(図2)
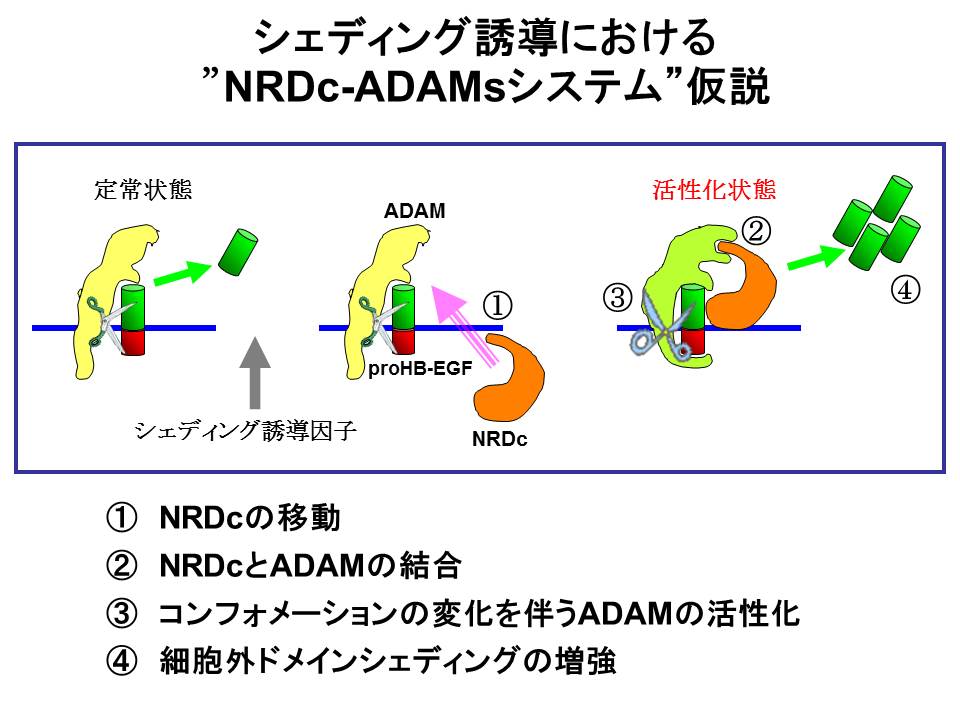
詳細は次項で述べますが、NRDCの遺伝子改変マウスを用いた検討から、NRDCがin vivo(生体内)においても、シェディング調節において重要な役割を果たしていることがわかってきました。
3) ナルディライジン(NRDC)の生物学的機能と病態生理学的意義
3)-1: 神経系発達におけるNRDCの役割
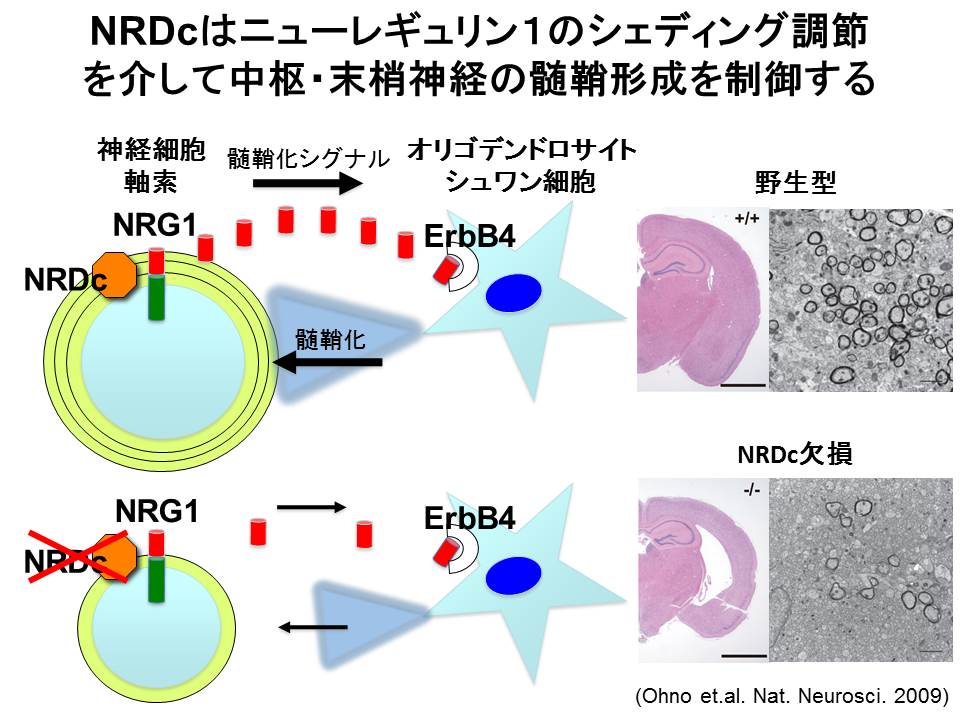
NRDC欠損マウス(神戸理研との共同開発)の作製は、私たちの研究の進展に大きく寄与しました。NRDC欠損マウスは出生時から明らかに体が小さく、生後の成長遅延も認めました。一見したところ、体が小さい以外は運動や摂食に大きな異常はなさそうでしたが、尾懸垂試験にて異常反射を認めたことから脳を観察したところ、大脳皮質の著明な菲薄化を認めました。詳しく調べてみると、神経細胞の数が減少しているのではなく、神経細胞体から伸びる軸索が細く、かつ軸索をくるんでいる髄鞘の厚みが薄いことがわかりました。この表現型は中枢神経系だけではなく末梢神経系でも認めたことから、NRDCが神経軸索の成熟と髄鞘形成を制御する重要な因子であることがわかりました。さらに、NRDCがニューレギュリン(EGFファミリー増殖因子)のシェディング調節を介して髄鞘形成を制御していることを明らかにしました(図3)。この神経系の表現型に関する論文は2009年にNature Neuroscience誌に報告し、多くの新聞でも取り上げて頂きました。
(図3)
3)-2: アルツハイマー病におけるNRDCの役割
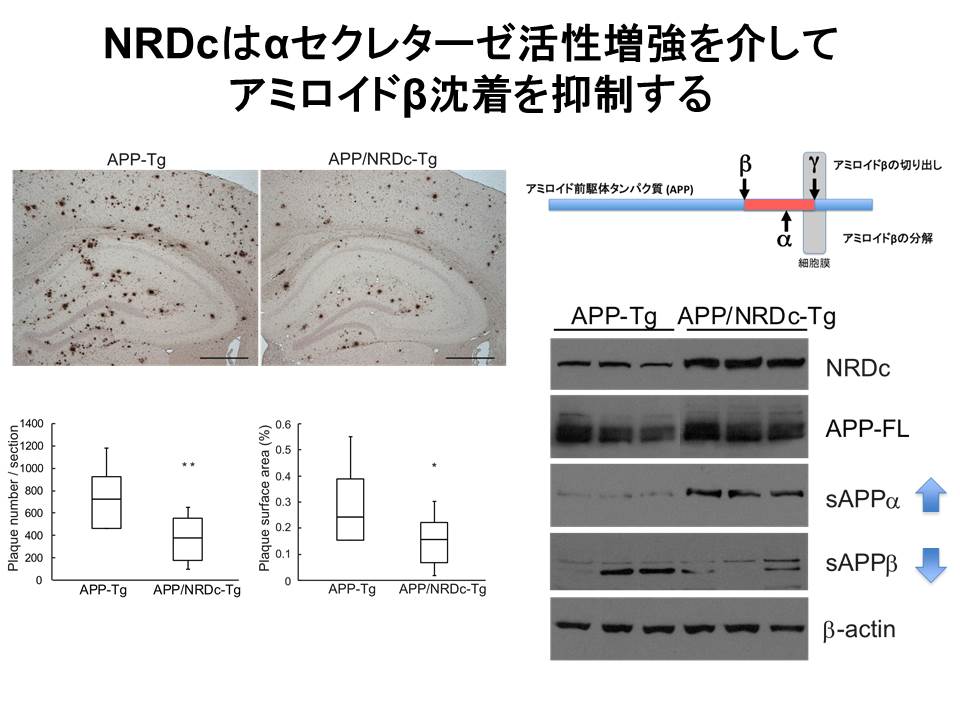
アミロイド前駆体タンパク質(APP)は異なるセクレターゼにより3カ所(α、βは細胞外ドメインシェディング、γは膜内切断)で切断されますが、アルツハイマー病の原因と考えられているアミロイドβは、βおよびγセクレターゼで切り出されます。一方、αセクレターゼはアミロイドβをその中程で切断することで、同産生を抑制すると考えられています(図4)。主たるαセクレターゼはADAMプロテアーゼであることが報告されていることから,我々はNRDCがαセクレターゼを活性化してアミロイドβの産生を抑制するのではないかと考え、細胞レベルで検討したところ、予想通りの結果が得られました(Hiraoka他J.Neurochem,2007)。次に、神経細胞特異的NRDCトランスジェニックマウスとアルツハイマー病モデルマウスとの交配実験を行ったところ、マウス個体レベルにおいても、NRDCがAPPのα切断増強を介してアミロイドβの産生を抑制することがわかりました(図4)。この結果は、NRDCがαセクレターゼ活性制御に重要な因子であり、アミロイドβ代謝制御、アルツハイマー病の病態生理に関与する可能性を示唆しました(Nurobiology
of Aging, 2014)。
(図4)
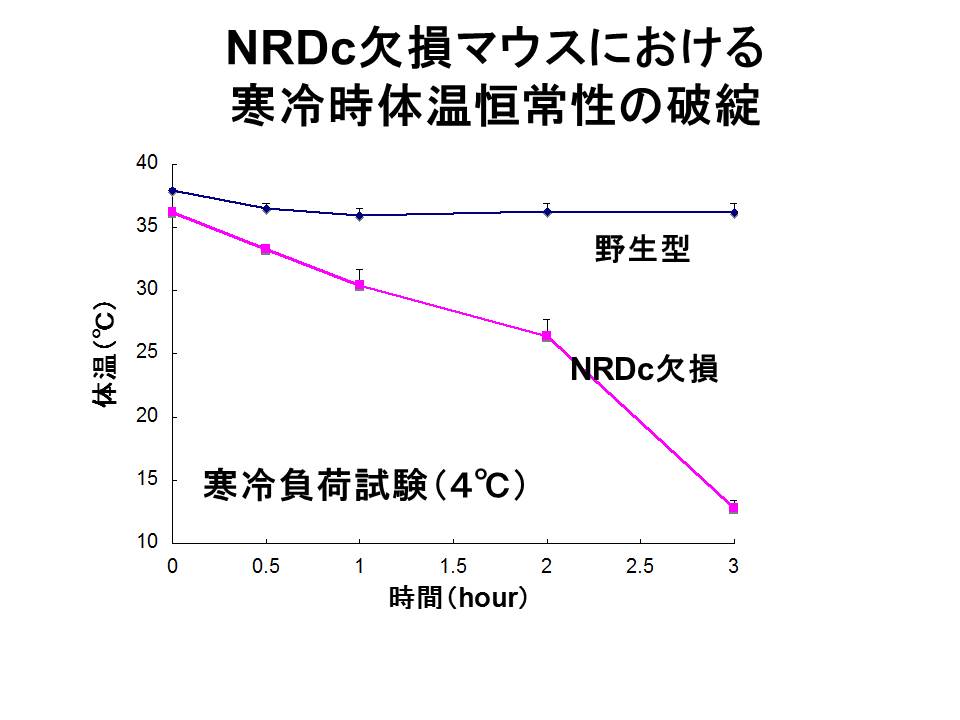
3)-3: 体温恒常性維持機構におけるNRDCの役割
NRDC欠損マウスは、野生型マウスと比較して常温で体温が1.5℃程度低く、低温環境(4℃)では体温が何と10℃台まで低下してしまいます(図5)。正常なマウスは、低温環境におかれると、熱放散を抑制するために皮膚血管を収縮させ、同時に主たる熱産生臓器である褐色脂肪細胞(brown
adipose tissue, BAT)における熱産生のスイッチをオンにします。NRDC欠損マウスのBATを解析した結果、意外なことに常温でのBAT熱産生はむしろ亢進していましたが、寒冷負荷による熱産生スイッチが入らないことがわかりました。
(図5)
詳細な検討から、NRDCは、①BATにおける熱産生、②皮膚血管における熱放散、さらに③中枢性体温セットポイント調節を、それぞれ独立に制御することで体温恒常性を維持していることがわかりました。
BATにおける熱産生は、ミトコンドリア脱共役タンパク質(UCP1)が担っており、UCP1の転写調節にはPGC1αという転写コアクチベーターが重要であることがわかっていました。我々は、核内でNRDCがUCP1の転写エンハンサー領域に存在し、PGC1αとの直接結合を介して、その転写コアクチベーター活性を制御し、その結果UCP1の発現を調節していることを明らかにしました。後述するように、2012年にNRDCがヒストンに結合して転写調節に関わることを報告しましたが、この仕事で、NRDCの核内機能がin
vivoでも重要な役割を果たしている可能性を示唆することができました。
この仕事はNature Communications誌に報告し、新聞でも報道されました。
3)-4: 糖尿病、メタボリック症候群におけるNRDCの役割
NRDC欠損マウスは、やせや血清中性脂肪の低下など、メタボリック症候群と対称的な表現型を呈します。一方驚いたことに、NRDC欠損マウスにグルコースを負荷しても、ほとんどインスリンが分泌されないことがわかりました。インスリンは膵β細胞で産生されるので、膵β細胞特異的にNRDCを欠損したマウスを用いて検討したところ、やはりグルコース応答性インスリン分泌は著しく減弱していました。以上から、膵β細胞に発現するNRDCがインスリン分泌制御に極めて重要な役割を果たすことが明らかになりました。
この仕事はDiabetes誌に報告し、新聞でも報道されました。
現在さらに、脂肪細胞や肝細胞に発現するNRDCがインスリン感受性の制御において果たす役割を追求しており、糖代謝におけるNRDCの統合的な役割を明らかにしようとしています。
3)-5: 循環動態制御におけるNRDCの役割
NRDC欠損マウスは、徐脈、低血圧を呈します。にもかかわらず、血清カテコラミンは高く、交感神経活動は亢進していることがわかりました。さらに、心臓における交感神経終末の分布パターンが大きく変化していることもわかり、NRDCが循環動態の恒常性維持にとても重要な役割を果たしていることがわかりました。心筋と交感神経それぞれ特異的にNRDCを欠損するマウスを作製することで、NRDCの役割をさらに明らかにしようとしています。
3)-6: 心疾患におけるNRDCの役割
我々が、三洋化成株式会社、ミクリ免疫研究所株式会社と共同開発した高感度NRDC測定系を用いて検討したところ、急性冠症候群やたこつぼ心筋症患者の血清NRDCが有意に上昇していることがわかりました。これらの疾患バイオマーカーとしての有用性を明らかにするとともに、心筋特異的NRDC強発現あるいは欠損マウスで心筋梗塞、心不全モデルを作製し、心疾患におけるNRDCの病態生理学的意義を明らかにしようとしています。
さらに、NRDCの血中濃度と血中自己抗体の解析を組み合わせることで、急性冠症候群(ACS)を、その発症前に予知することができないか、という課題に取り組んでいます。初回の抗原曝露後の抗体産生には2週間以上かかることから、ACS発症後早期に上昇している自己抗体は、発症前から出現していたはずであり、自己抗体をスクリーニングすれば予知マーカー探索が可能になると考えられたからです。本プロジェクト(千葉大学遺伝子生化学 日和佐先生との共同研究)は厚生労働省科学研究費に採択していただき、実地臨床で役立つ予知マーカーの開発に取り組んでいます。
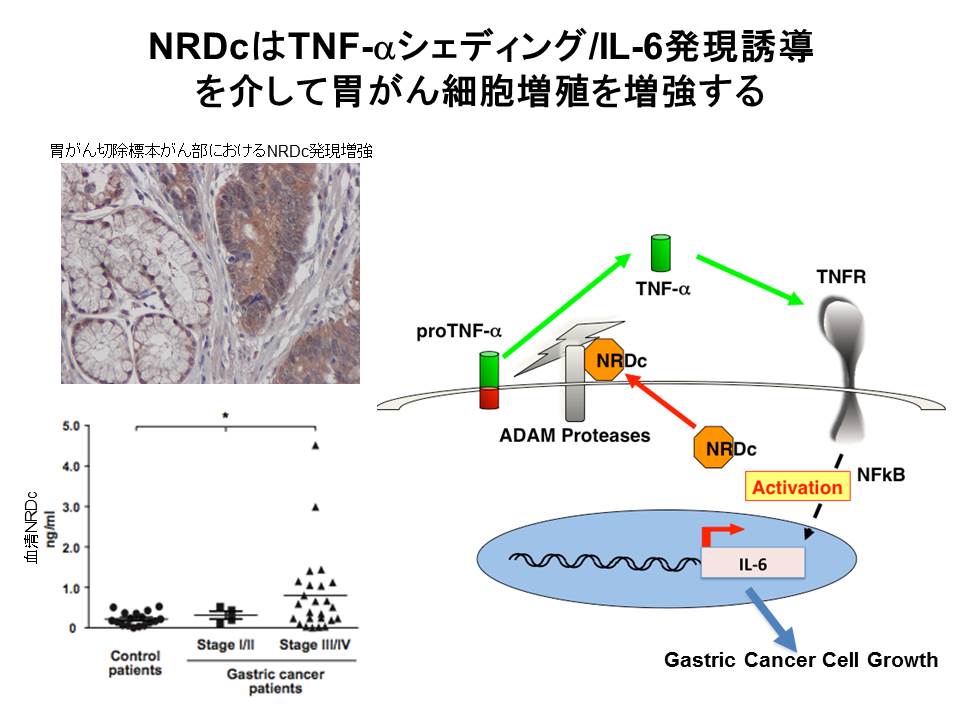
3)-7: がんにおけるNRDCの役割
前述した高感度測定系で検討したところ、NRDCはいくつかのがん患者血清中で有意に上昇していました。消化器内科の妹尾先生グループとの共同研究にて、NRDCの発現が胃がん病変部(切除標本)で増加していること、胃がん細胞株でNRDCの発現を抑制すると細胞増殖が抑制されること、を明らかにしました。分子メカニズムとしては、NRDCが炎症性サイトカインTNF-αのシェディング増強を介して、その下流のインターロイキン6発現を誘導し、胃がん進展に寄与していることが示唆されました(図6)(Kanda
et.al. EMBO Mol Med, 2012)。
(図6)
その後、NRDC欠損マウスが複数のがんモデルにおいて著明な発がん抵抗性を有することが明らかになっています。学内では、消化器内科、肝胆膵移植外科、泌尿器科、乳腺外科の先生方にもご協力いただき、それぞれの領域におけるNRDCの重要性を検証しようとしています。本プロジェクトは文部科学省の次世代がん研究シーズ戦略的育成プログラムにも採択され、NRDCを標的とする新たな抗がん療法開発を目指しています。
3)-8: 炎症性疾患におけるNRDCの役割
がんや生活習慣病の病態基盤として、慢性炎症が重要な役割を果たすことが明らかになってきています。これまで述べてきたように、NRDCは炎症性サイトカインTNFαや増殖因子HB-EGFの活性化に関与しており、炎症制御に何らかの役割を果たす可能性があります。
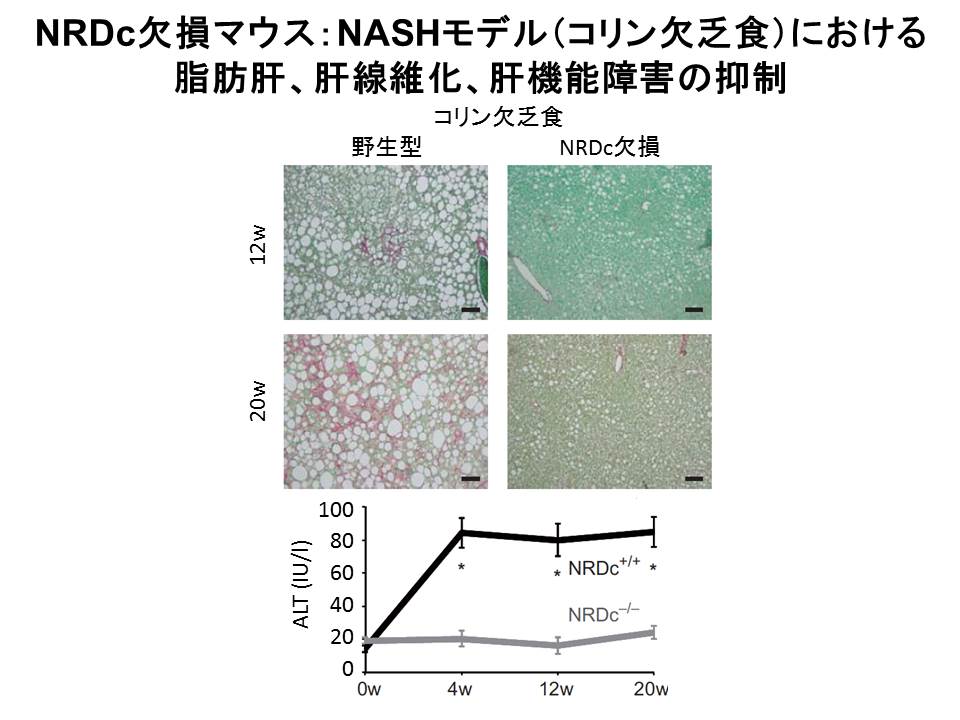
消化器内科の妹尾先生グループとの共同研究において、マウス非アルコール性脂肪性肝炎(NASH)モデル(コリン欠乏食、高脂肪食)を用い、NRDC欠損マウスは野生型マウスと比較して、脂肪肝になりにくく、肝機能障害、肝線維化の程度は軽度であることを明らかにしました(図7)(PLOS
ONE, 2014)。
(図7)
その他、動脈硬化、慢性関節リウマチ(整形外科 伊藤先生グループとの共同研究)におけるNRDCの役割についても研究を進めています。
4) ナルディライジンの多機能性
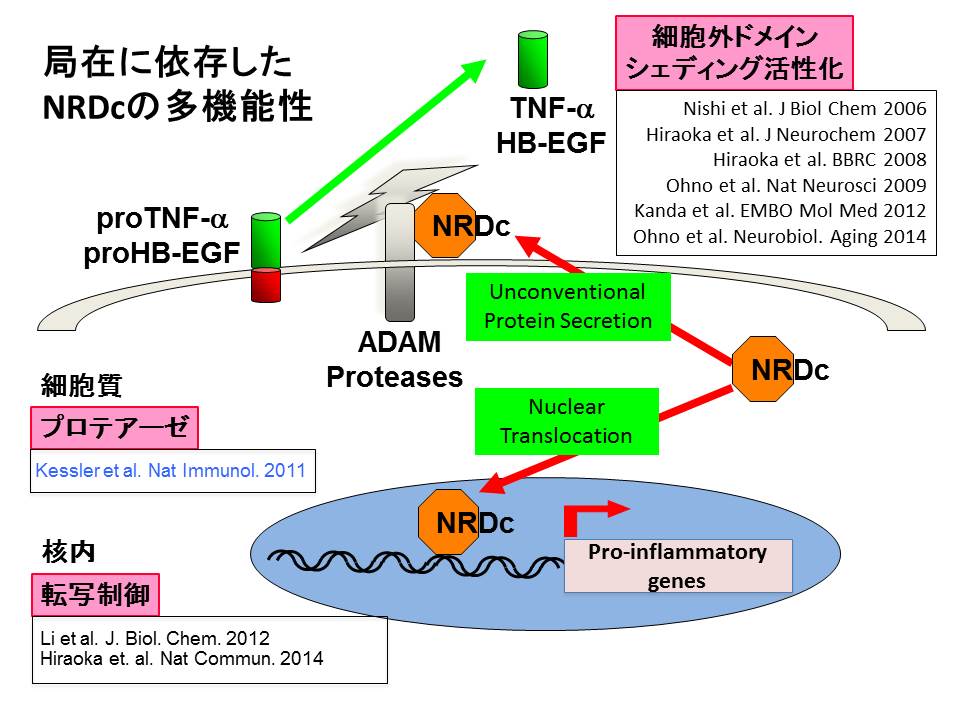
前項でざっと掲げたように、NRDC欠損マウスは成長障害、軸索・髄鞘低形成、低体温、徐脈、低血圧、糖脂質代謝異常、がん・炎症抵抗性、など極めて広範な表現型を呈しました。これまで、表現型から類推される分子メカニズムにおけるNRDCの役割をひとつひとつ確認するという作業を行い、例えば髄鞘形成におけるニューレギュリンのシェディング調節や、BAT熱産生におけるUCP1発現調節におけるNRDCの重要性を明らかにしました。
ノックアウトマウス作製前にわかっていた細胞外機能(細胞外ドメインシェディング活性化機能)については、ニューレギュリン、APP、TNFαなどのシェディングを介してin
vivoでも重要な機能を果たしていることがわかりました。
一方、NRDCが核と細胞質をシャトリングすることはずっと前からわかっていたのですが、その核内における機能は、ふたつの独立した実験系から漸くその一部が明らかになってきました。
ひとつは、East China Normal UniversityのWong博士との共同研究で、NRDCがヒストンにその修飾依存的に結合すること、すなわちH3,
H3K4me1-3, H3K9me1-3, H3K27me1-3の中で,H3K4me2に特異的に結合することがわかりました。さらに核内でNRDCと複合体を形成するタンパク質を解析したところ,NCoR,
HDAC3, SMRTが同定され、NRDCがNCoR/SMRTコリプレッサー複合体に含まれている可能性が示唆されました。実際に複数の遺伝子プロモーター上にNRDCが存在して、その転写を制御していることがわかりました(Lin他
JBC 2012)。
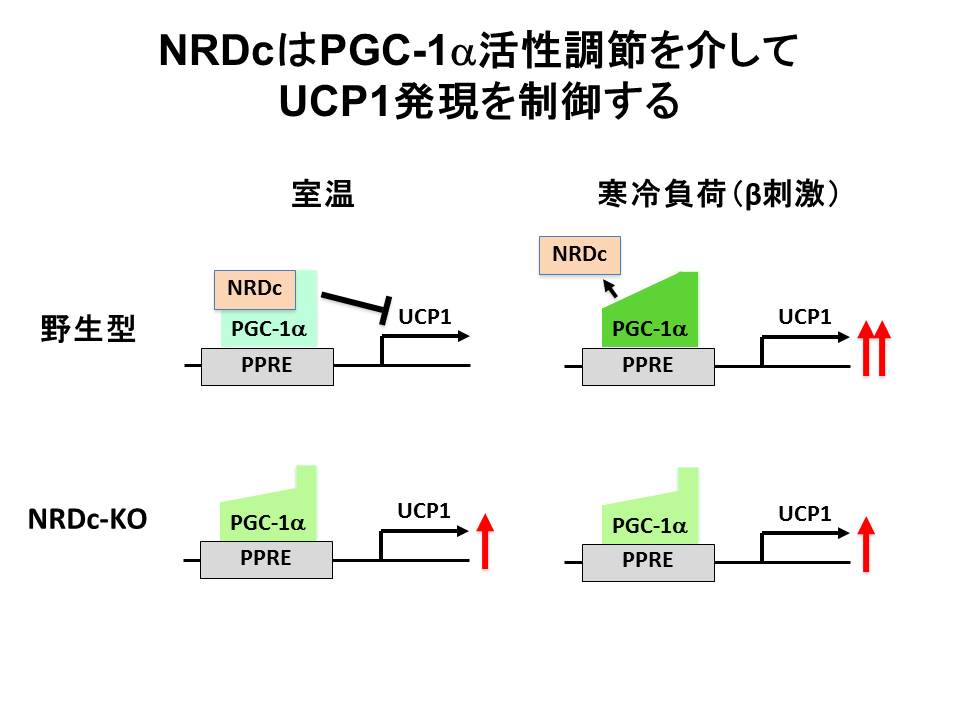
もうひとつは、NRDC欠損マウスの低体温と寒冷不耐性の表現型に端を発した系です。詳細な解析から,NRDC欠損マウスが体温を保持できないのは、BATにおける熱産生が常温ですでにピークに達しており、寒冷負荷時に亢進しないためであることがわかりました。先述したとおりBATにおける適応熱産生は、ミトコンドリア脱共役タンパク質(UCP1)に依存しており、UCP1の発現は、転写コアクチベーターPGC-1αの制御を受けています。NRDCはUCP1の転写エンハンサー領域に存在しPGC-1αと共在し、常温ではPGC-1αの転写コアクチベーター活性を抑制しています。低温では、何らかの機序によりNRDCがPGC-1αと解離し、UCP1発現上昇、熱産生誘導につながります。一方NRDCが欠損していると、低温で起こるべき誘導がかからず、体温維持に必要な追加の熱産生が起こらないことが示唆されました(図8)(Nat.
Commun. 2014)。
(図8)
さらに膵β細胞においては、核内NRDCがislet1という転写因子と協調して、MafAというβ細胞特異的転写因子の発現を調節していることがわかりました。NRDCは、この経路(islet1-MafA
経路)を介して、グルコース応答性インスリン分泌の制御に重要な役割を果たしています(Diabetes 2016)。
NRDC自身は明らかなDNA結合ドメインを有しませんが、以上の結果からNRDCが状況に応じて様々な転写因子、転写コアクチベーター、コリプレッサーと複合体を形成し、転写コレギュレーターとして転写を調節していると考えられます。
以上からNRDCが細胞外と核内で全く異なる機能を有することが明らかになりました。さらに他のグループからNRDCのプロテアーゼ機能が、細胞傷害性T細胞における抗原プロセッシングに必要であることが報告されており(Nat
Immunol 2011)、NRDCが局在依存性に異なる機能を持つ多機能タンパク質であることが示唆されています(図9)。
(図9)
今後は、臓器特異的NRDC欠損マウスや、特定の機能を欠損する変異型NRDCノックインマウスを用いて、「どの臓器に発現するNRDCの、どの機能が重要なのか」を明らかにしていきたいと考えています。
5) おわりに
NRDC欠損マウスは、低体温、徐脈、低血圧、低インシュリン血症などを呈するにもかかわらず、正常なネガティブフィードバックが機能せず、あたかも異なるセットポイントで平衡を保ちつつ生命を維持しているように見えます。それぞれのセットポイントがいかに決定されているか、その分子機構はほとんど明らかになっていません。人為的にセットポイントを変化させることが可能かどうか、それを疾患治療に応用することができないか、など考えながら研究を進めていきたいと思っています。
ゲノムプロジェクトの成果から、ヒトを初めとする高等生物において、タンパク質をコードする遺伝子の数が予想以上に少ないことが明らかになりました。高等生物の複雑な生命現象が、少ない数のタンパク質でいかに制御されているかについて、ひとつのタンパク質分子が複数の異なる機能を発揮することで貢献しているという説明は、理に適っているものの充分には検証されていません。なぜならタンパク質の機能、特にin
vivoにおける生物学的機能は、その局在解析、遺伝子改変動物の作製と表現型解析、結合タンパク質の同定など、非常に手間のかかる多くのステップを経て初めて明らかにすることができるからです。特に、特徴的な機能ドメインを持たないタンパク質の機能、さらに機能ドメインから推測できる機能以外を明らかにするためには、多くの労力が必要です。私たちは、多くの共同研究者に支えられつつ、NRDCに関するマテリアル、例えばNRDCに対する多種類の抗体、高感度測定系、遺伝子改変マウスなど、全てを自分たちの研究室で開発し、解析を続けてきました。最近少しずつですが、世界中の研究室からマテリアルのリクエストが増えてきたことを嬉しく思っています。今後さらに多くの方々と協力しながら、このユニークで魅力ある分子の機能を明らかにし、その中で疾患の新たな診断法、治療法開発につなげることができれば、と考えています。
